|
Elementos Minerais Essenciais ao Desenvolvimento das Plantas A Meireles(a) e A Fernandes(b) (a)Aluno 30358, Fisiologia Vegetal – Eng. Agronómica UTAD, 2º ano (b)Aluna 30359, Fisiologia Vegetal – Eng. Agronómica UTAD, 2º ano
Resumo Com este trabalho prático pretendeu-se verificar o comportamento das plantas perante a ausência de determinados minerais essenciais ao seu desenvolvimento. Para isso, procedeu-se à determinação do comprimento máximo de caules e raízes, posteriormente ao registo dos sistemas de deficiência e também ao registo dos sintomas de toxicidade associados à salinidade elevada. De salientar que também se registou as variações de pH obtidas em três frascos, esta análise foi realizada durante três semanas com intervalos semanais. Toda esta experiência foi realizada em meio hidropónico com plantas de milho( Zea mays). Palavras-chave: plantas, minerais, deficiência, salinidade, pH. 1-Introdução O trabalho prático sobre os elementos minerais essenciais ao desenvolvimento da planta pretende analisar o comportamento das plantas quando confrontadas com a falta de certos elementos minerais essenciais ao seu desenvolvimento. A aquisição de compostos químicos realizada por um organismo para suprir o seu metabolismo é chamada de nutrição. O metabolismo compreende os processos pelos quais os compostos químicos (nutrientes) são utilizados para o crescimento e manutenção da planta. Os nutrientes podem, principalmente, ser convertidos em material celular ou ser usados como fonte de energia. As plantas, geralmente autotróficas, não dependem do fornecimento de compostos ricos em energia produzidos por outros organismos, como, ocorre nos seres heterotróficos. Uma separação didáctica tradicional organiza a nutrição das plantas está organizada em duas formas, a forma orgânica e a forma inorgânica. A orgânica refere-se à aquisição de carbono, oxigénio e hidrogénio, provenientes da atmosfera e da água, via fotossíntese e a nutrição inorgânica, também chamada mineral, refere-se aos restantes elementos que geralmente são absorvidos do solo. Só foi possível um aumento da eficiência agrícola, devido a uma melhor compreensão dos mecanismos de nutrição mineral das plantas. Devido a este conhecimento superior da nutrição mineral, houve consequentemente um aumento da produtividade das culturas agrícolas, independentemente da evolução tecnológica a nível das máquinas agrícolas, uma vez que as máquinas apenas ampliam capacidade de exploração mas não contribuem tanto para o aumento da produção vegetal por unidade de área. Aristóteles (384-322 AC) acreditava que as plantas se alimentavam do húmus do solo. Os gregos incluíam a Botânica na Filosofia e grande parte de suas conclusões não eram baseadas em experimentação, mas sim em associação antropomórfica. Posteriormente pesquisadores como, por exemplo, Van Helmont (1577-1644) postularam que a planta só retiraria água do solo. Esta interpretação errónea foi induzida pelo facto de que cerca de 95% da matéria seca das plantas vem da nutrição orgânica e somente o restante são minerais do solo. Foi por este motivo que a planta observada por Van Helmont durante 5 anos pareceu não reduzir a quantidade de solo do seu vaso. Um dos primeiros avanços nesta área do conhecimento foi gerado pelo trabalho de N.T. de Saussure (1767-1845) comprovando que as plantas absorvem os elementos do solo com taxas diferenciadas, não funcionando apenas como uma esponja. Saussure postulou ainda que nem todos os elementos presentes na planta são essenciais. C.S. Sprengel (1787-1859) introduziu a ideia de que o solo pode ser improdutivo se for deficiente em apenas um dos elementos essenciais às plantas. Em 1860 Julius Sachs realizou a primeira experiência hidropónica conseguindo que as plantas produzissem recebendo somente nutrientes minerais em solução e sem solo. No final do século XIX a adubação com N-P-K (nitrogénio, fósforo e potássio) já estava estabelecida na Europa. A hidroponia apresenta as seguintas vantagens: . Produção em pequenas áreas; . Utilização de pequenos volumes de água e controlo da qualidade da mesma; . Redução drástica do uso de quimicos; . Redução do nº de operações durante o ciclo de vida da cultura; . Antecipação da colheita; . Qualidade superior do produto colhido; . Produção fora de época; . Riscos reduzidos de perda da produção devido a factores climáticos; . Rápido retorno económico.
Figura 1- Produção intensiva em meio hidropónico 2- Material e Métodos Material: - Frascos - Água destilada - Placa de esferovite com três buracos - Três plantas de milho - Medidor de pH Método: 1) Etiquetar 11 frascos de vidro, como se segue: FeEDTA completo, FeCl3 completo, -Ca; -K, -S, -Mg, -N, -P, -Fe, -Micronutrientes, +salino. 2) Encher os frascos até cerca de 2/3 com água destilada (solução 100mM NaCl no tratamento +salino); 3) Juntar os volumes das soluções stock indicadas no quadro em anexo; 4) Agitar. 5) Encher todos os frascos até cerca de 1 cm do bordo. 6) Introduzir 3 plantas de milho nos suportes de esferovite; 7) Aconchegar com algodão, mas evitar o contacto deste com a solução. 8) Antes de colocar os frascos na câmara de crescimento, determinar o pH das soluções Fe EDTA completo, –N e + salino; Repetir a intervalos semanais. 9) Adicionar água destilada (ou a solução 100mM NaCl), sempre que necessário, para manter o nível inicial das soluções; Fazer o arejamento regular. 10) Remover uma plântula de cada frasco, após uma semana e ao fim de 3 semanas. 3- Resultados Quadro 1- Valores Ph obtidos para os frascos sem FeEDTA, N e Salino(+) (Grupo 1)
Quadro 2- Valores
Ph obtidos para os frascos sem FeEDTA, N e Salino(+)
Quadro 3-
Comprimentos de caules e raízes obtidos nos diferentes frascos
Quadro 4-
Comprimentos de caules e raízes obtidos nos diferentes frascos
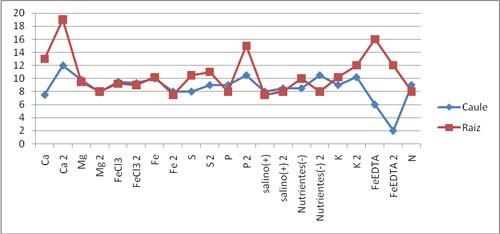
Figura 2- Comprimento do caule e da raiz das plantas em diferentes meios (Grupo1)
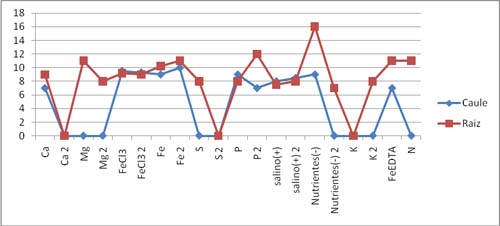
Figura 3- Comprimento do caule e da raiz das plantas em diferentes meios (Grupo2) 4- Discussão de Resultados Com a elaboração deste trabalho pretendia-mos avaliar o crescimento e a resistência das plantas de milho em relação á carência de um tipo de nutriente essencial para o seu desenvolvimento. Os resultados obtidos não foram os esperados, devido principalmente as baixas temperaturas que se fazem sentir em Vila Real nesta época do ano, o que faz com que se dê uma forte inibição do crescimento das plantas. Ao longo da passagem dos dias da experiência fomos verificando o estado das plantas e medindo o ph dos frascos que continham, FeEDTA, N, Salino(+), e verificou-se uma diminuição do ph. De salientar que no grupo 2, houve alguns erros, na medida em que há variações de pH nestes três frascos, e não uma diminuição do pH há medida que os dias iam avançando. O pH deveria diminuir em todos os frascos em que se procedeu à sua medição devido à oxigenação do meio, por nós realizada, no decorrer da experiência. Os sintomas de deficiência mineral que deviam ter sido detectados são: - Nas folhas mais novas: . Clorose geral progredindo de verde-claro para amarelo; . crescimento atrofiado; folhas necróticas podendo secar completamente devido à falta de azoto(N); . Clorose nas margens e extremidades das folhas, podendo progredir para necroses; folhas frágeis e encurvadas para cima devido à falta de magnésio(MG); . Margens das folhas acastanhadas ou manchadas e recurvadas para baixo devido à falta de potássio(K). . Acumulação de antocianinas nas folhas dando-lhe uma coloração verde-azulada ou vermelho-púrpura; as folhas inferiores podem tornar-se amarelas devido à falta de fósforo(P). - Nas folhas mais velhas: . Áreas amarelas ou esbranquiçadas entre as nervuras; inicialmente as nervuras são verdes tornando-se cloróticas em condições de deficiência mais severa devido à falta de ferro(Fe). . Folhas verde-claras, progredindo para amarelecimento; tecidos interveinais com cor clara devido à falta de enxofre(S). - Gomos Terminais: . Parcial ou total ausência de regiões meristemáticas; extremidades e margens das folhas necróticas devido à falta de cálcio(Ca).
Figura 4-Pé de Milho com deficiência de azoto
Figura 5-Pé de Milho com deficiência de Cálcio
Figura 6-Pé de Milho com deficiência de Ferro
Figura 7-Folha de Milho com deficiência de Potássio
Figura 8-Folha de Milho com deficiência de Magnésio
Figura 9-Folha de Milho com deficiência de Magnésio
Figura 10-Folha de Milho com deficiência de Enxofre 5- Conclusão Com a realização deste trabalho chegamos á conclusão que existem nutrientes extremamente essências para o desenvolvimento das plantas. Quando existe uma carência de um qualquer nutriente essencial, este vai afectar o rendimento final da cultura e mesmo em muitos casos dá-se a morte da mesma, tendo deste modo uma enorme quebra da produção e consequentemente um elevado prejuízo económico. Quanto ao sucesso da experiência podemos dizer que não foi bem conseguida devido principalmente ao factor temperatura, visto que em Vila Real fazem-se sentir temperaturas muito baixas nesta altura do ano, o que inibe em muito o crescimento das plantas e assim, a nossa posterior identificação dos sintomas da carência mineral. Chegamos também á conclusão que a produção de plantas em substrato liquido, isto é, em meio hidropónico é extremamente vantajosa principalmente porque nos permite ter uma produção fora de época e pode também ser efectuada em espaços menores. 6- Bibliografia [Web] [http://fisiologiavegetal.homestead.com/files/Nutricao_mineral_de_plantas.htm](em 17/12/2009) [Web] [http://www.lumaplastic.com.br/como_funciona_hidroponia.htm](em 17/12/2009)
Os efeitos das micorrizas nativas e não nativas no crescimento e nutrição da “Pinot Noir”( Vitis vinifera L.) em dois solos com níveis distintos de fósforo A.Meireles a a1Aluno nº30358, Curso de Engenharia Agronómica da UTAD, afonsoportista@msn.com Introdução Este trabalho tem como objectivo mostrar a influência que as micorrizas podem exercer sobre o crescimento e nutrição na vinha, mais precisamente na casta “Pinot noir”. Esta experiência foi realizada em dois tipos de solos, Chehalis e Jory. Chehalis é um solo característico do fundo do vale Oregon e o solo de Jory é um solo característico da encosta do vale Oregon. Estudos previamente realizados na área da vinha, provaram que a casta “ Pinot Noir” necessitava das micorrizas(AMF- Fungo Micorrizico Arbustivo) para a promoção de um crescimento e desenvolvimento normal. Os estudos realizados demonstraram que a presença de AMF favorece o aumento da absorção de fósforo por parte da videira, não se sabendo qual o papel dos AMF na absorção de outros nutrientes por parte da planta. Verificou-se neste estudo um contraste entre os dois solos, uma vez que os solos da encosta vermelha (Ultisols of Jory) localizados na zona de Oregon são caracterizados por possuírem uma alta acidez, baixa fertilidade, serem altamente intemperados e com baixa concentração de fósforo (10 mg kg-1), e por outro lado, o outro solo utilizado no estudo pertence à outra região vinhateira localizada no fundo do vale, abaixo das encostas de Oregon (Chehalis), sendo que estes solos, devido à sua localização geográfica e por seres solos aluviais tem uma maior fertilidade de que os solos da encosta vermelha. Em ambos os solos, as videiras, têm mostrado teores de colonização por parte de AMF altos, apesar da diferença de fertilidade existente entre estes dois solos, principalmente a nível de fósforo e cálcio, sendo maior esta concentração nos solos do vale. Confirmou-se que para determinado solo, os fungos nativos são mais eficazes do que os fungos não nativos, devido à sua adaptação aos factores edáficos da região. O objectivo principal deste estudo é compreender se o crescimento da vinha situada no solo da encosta tem uma maior dependência de AMF para a obtenção de nutrientes, nomeadamente a concentração de fósforo, comparando o crescimento da vinha no solo do vale. Aquando da obtenção dos resultados foi comparado o comportamento das plantes que possuíam as micorrizas com as plantas que não possuíam as micorrizas. Materiais e Métodos Para a realização deste estudo, foram colhidas amostras de ambos os solos, sendo que estas amostras foram colhidas de 0 a 30 cm de profundidade, de salientar que ambos os solos foram colhidos pela Oregon State University. Estas amostras foram secadas e esterilizadas a 150ºC durante 48horas e na amostra de solo Jory (JY) foi adicionado calcário dolomítico para que assim este apresentasse um pH semelhante ao do solo Chehalis (CH), devido a esterilização a que são sujeitos, estes solos são previamente armazenados durante 2 a 4 meses. Posteriormente foi utilizado um ramo da poda, com um ano de idade, colhida pela Woodhall Research Vineyard, sendo depois armazenada a uma humidade e temperatura de 4ºC por 3 meses para mais tarde produzir três ramos que seriam usados na experiência. O trabalho divide-se em duas experiências: . Experiência 1: teste de mistura de AMF nos dois solos; . Experiência 2: teste do nativo versus não nativo G. mossae nos dois solos. Experiência 1 Seis replicações foram incluídas em cada tratamento para um total de 24 plantas envasadas (unidades experimentais). Dentro de cada tipo de solo, metade dos vasos receberam a mistura de AMF, e a outra metade servirá de grupo de controlo em que não recebe a mistura de AMF. Os AMF’s inoculados são: . Scutellospora calospara; . G. mossae; . Glomus sp. Todos os fungos tiveram um isolamento prévio do solo Jory da Woodhall Research Vineyard, sendo posteriormente cada espécie de fungo isolada para serem propagadas em esporos e novamente cultivadas, em recipientes com solo arenoso, muito fraco em concentrações de fósforo, e com a presença de Sorghum bicolor L. Para a promoção de um crescimento dos AMF’s foi depositado nos seus tratamentos solo inoculado contendo raízes, plantas, esporos e hifas. Depois dos tratamentos foi depositado em cada pote (vaso) um extracto microbial, correspondente à concentração microbial nos dois tipos de solos quando não estavam tratados, e concentração de AMF inoculado. Foi realizada tal operação de modo que os solos tratados ficassem com concentrações microbianas e de AMF o mais semelhante possível às concentrações que possuem no seu ambiente natural, tendo sido o extracto obtido através por um crivo de 38μm. As plantas que posteriormente iriam ser utilizados na experiência cresceram em estufa, sendo que a temperatura foi ajustada de forma a corresponder à temperatura no exterior (14-20ºC/20-30ºC) para noite e dia respectivamente, não apresentarando stress hídrico. Também foram fertilizadas com 400ml de solução de Hoagland (Hoagland e Arnon, 1950) uma vez de duas em duas semanas sendo, a solução de Hoagland uma solução nutritiva para plantas em hidropúnia. Experiência 2 Seis replicações foram incluídas em cada tratamento para um total de 36 unidades experimentais (plantas envasadas). Em cada tipo de solo, um terço dos vasos receberam o G.mossae inoculado, isolado do Chehalis solo, o outro terço recebeu o G.mossae incoculado tendo sido isolado do solo Jory e outro terço não recebeu os AMF sendo denominados de vasos do controlo. Cada pote de micorrizas recebeu 300 propágulos infectados do fungo G.mossae. Mais tarde o rebento principal da planta atingiu uma altura característica (cerca de 140 cm), como resposta a este facto, os rebentos laterais começaram a crescer. Resultados Verificou-se na experiência 1, que no solo CH, o crescimento dos rebentos de “Pinot noir” das plantas sem AMF (grupo de controlo) apresentaram um crescimento significativamente maior do que os rebentos que possuíam AMF (incluídos nos tratamentos), dado que estes resultados foram obtidos 57 dias depois da plantação. Oitenta e quatro dias após a plantação, o crescimento das plantas de controlo e das plantas de tratamento já tinham valores muito próximos acabando por mais tarde ter um crescimento muito semelhante. Por sua vez, no solo Jory, as plantas com tratamento tiveram um crescimento muito similar em relação às plantas do grupo de controlo, isto nos primeiros 57 dias, contudo, 84 dias depois da sua plantação os rebentos das plantas com tratamento apresentavam um crescimento maior em relação ao grupo de controlo, deve-se no entanto salientar que 84 dias após a plantação, o grupo de controlo no solo Jory tem tendência a parar o crescimento, e já o grupo com tratamento tem um crescimentos exponencial adquirindo valores tão altos como os valores adquiridos pelos rebentos do solo Chehalis. Houve uma maior retenção de biomassa no solo Jory, sendo que as plantas em que AMF estava presente reteram três vezes mais biomassa do que as plantas do grupo de controlo, por outro lado no solo Chehalis, não foram encontradas diferenças significativas entre os dois grupos( Com e sem AMF). Não se obteve qualquer diferença entre os dois grupos nos dois tipos de solos em relação à razão rebento/raiz. A nível de comprimento radicular, o grupo de controlo, no solo Chehalis, apresentou um crescimento muito superior em relação às plantas pertencentes ao grupo com presença de AMF, por outro lado no solo Jory as plantas com AMF apresentaram um crescimento radicular superior ao grupo de controlo. A percentagem de colonização de AMF, foi superior em todos os aspectos nas plantas que se encontravam no solo Jory. Quanto à nutrição das plantas, verifcou-se que quase todas as concentrações de nutrientes sofreram alterações com a presença de AMF, exceptuando o zinco (Zn), o cobre (Cu) e o cálcio (Ca). No solo Jory, estando presentes AMF, as concentrações de nutrientes diminuiram com excepção do potássio (K) e do fósforo (P)(aumentaram), já no solo Chehalis, verificou-se que para as mesmas condições, relativamente à presença de AMF, o manganês (Mn) e o azoto (N) foram os dois nutrientes que obtiveram um maior grau de crescimento da sua concentração, e por outro lado as concentrações de Ferro (Fe), e de Boro (B) diminuiram com a presença dos fungos. As concentrações de nutrientes contidas nas plantas são inversas às concentrações de nutrientes existentes nos vasos. Assim, nas plantas com a presença de AMF no solo Jory obtiveram-se altas concentrações de todos os nutrientes. Por outro lado,as plantas que se encontravam no solo Chehalis obtiveram altas concentrações de macronutrientes, já as concentrações de micronutrientes baixaram com a presença de AMF. Quando comparadas as duas experiências, verificou-se que o crescimento final dos rebentos do Pinot noir foi aproximadamente o triplo na experiência 2 face à experiência 1, exceptuando as plantas de controlo do solo Jory. De salientar que a adição do fungo G.mossae, inoculado de cada solo, melhorou ligeiramente o crescimento dos rebentos no solo Chegalis, 51 e 62 dias após a plantação, mesmo assim as plantas com G.mossae e as plantas do grupo de controlo obtiveram valores muito semelhantes 90 dias após a plantação. Enquanto que no solo Jory, houve um aumento exponencial no que toca ao crescimento das vinhas onde se encontrava presente G.mossae, no solo Chehalis, o grupo de controlo não apresentou um crescimento significativo, sendo que não foi apresentado qualquer crescimento depois de 35 dias após a plantação. Quanto à acumulação de matéria seca nas videiras com a presença do fungo não se notou grande variação no crescimento das videiras no solo Chehalis. No entanto, a massa dos rebentos no solo Chehalis foi três vezes superior na experiência 2 do que na experiência 1. No solo Jory existiu um aumento pouco significativo com a presença do fungo, existindo um aumento muito superior se comparado com os resultados obtidos na experiência 1. Pôde-se também constatar que as concentrações dos nutrientes nas plantas com inoculação de G.mossae foram afectadas, à excepção das concentrações de Fe. As concentrações de nutrientes retidas no solo Chehalis, pelos diferentes grupos são muito semelhantes, por outro lado, no solo Jory, as concentrações de nutrientes aumentaram significativamente, quando comparadas com as obtidas no grupo de controlo no mesmo solo. Os grupos que possuíam os dois tipos de G.mossae inoculados no solo Chehalis, apresentaram valores superiores ou muito semelhantes, no caso da retenção de N, aos valores retidos pelo grupo de controlo. Já no solo Jory, os grupos onde o fungo inoculado estava presente, obtiveram sempre concentrações de nutrientes superiores aquelas que eram apresentadas pelo grupo de controlo.No solo Chehalis, a captação de Cu, foi superior no grupo que possuía o G.mossae nativo do solo, já na captação de P, o grupo que possuía o fungo não nativo obteve maiores concentrações. No solo Jory, os grupos que apresentavam a inoculação do fungo não nativo, que pertencia ao solo Chehalis conseguiram obter maiores concentrações de Cu e P, e apresentar valores muito semelhantes na concentração de N em relação ao grupo que continha o fungo nativo. Conclusão Pode-se concluir que o solo Jory, é pobre em P,e neste caso as videiras dependem intrinsecamente da presença de AMF para captação do mesmo, ainda que também, em menor escala, existisse um aumento na captação dos outros nutrientes, esta captação pode também ocorrer devido a uma maior captação de fósforo por parte da planta. Outro facto, que deve ser salientado, prende-se com a não deficiência em concentrações de P, descritas nas videiras, apresentadas pelo grupo de controlo, mesmo assim, ficou provado que as videiras precisam da presença de MAF para o seu crescimento, mesmo que se encontrem em solos secos, mesmo que estes solos apresentem altos níveis de concentração de P. No solo Chehalis, devido à sua fertilidade, a presença ou ausência de AMF não afecta os níveis de concentração, uma vez que as acumulações mais altas de S, Zn, ou Cu foram induzidas pelos altos níveis de P acumulado, que beneficiaram um bom crescimento da planta. Por fim, pode-se afirmar que os AMF nativos não são necessariamente os mais bem adaptados a um tipo de solo na promoção e retenção de nutrientes e que a capacidade de retenção de Cu pela planta varia em diferentes populações do mesmo fungo podendo fornecer assim um desenvolvimento de uma função específica para inoculação em uso prático.
Outros Trabalhos Relacionados
|
|