
O teu país
Lorem ipsum dolor sit amet, consectetur adipiscing elit, sed do eiusmod
Lorem ipsum dolor sit amet, consectetur adipiscing elit, sed do eiusmod

Todos os trabalhos publicados foram gentilmente enviados por estudantes – se também quiseres contribuir para apoiar o nosso portal faz como o(a) Cláudia Fernandes e envia também os teus trabalhos, resumos e apontamentos para o nosso mail: geral@notapositiva.com.
Resumo/Apontamentos sobre o Património Genético e Alterações do Material Genético, realizado no âmbito da disciplina de Biologia (12º ano).
A genética é uma ciência jovem, iniciada nos primórdios do séc. XX, altura em que foram redescobertas as leis 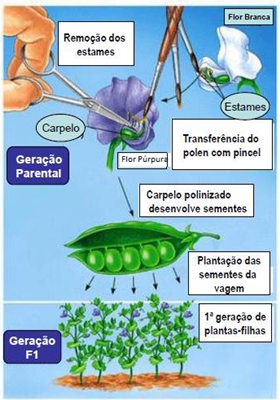 da hereditariedade, estabelecidas por Gregor Mendel.
da hereditariedade, estabelecidas por Gregor Mendel.
Os estudos que Mendel desenvolveu sobre a transmissão das características foram realizados em ervilheiras. Mendel escolheu estas plantas porque elas reuniam um conjunto de características que se adequavam ao trabalho que ele pretendia desenvolver:
Para se realizar polinização cruzada, removem-se os estames numa fase em que ainda não são capazes de produzir grãos de pólen e, posteriormente, fertiliza-se essa planta com o pólen proveniente de outra planta escolhida.
Mendel teve o cuidado de utilizar linhas puras, isto é, plantas que, quando autopolinizadas, originam uma descendência igual entre si e igual aos seus progenitores (relativamente a uma determinada característica). Para obter linhas puras, Mendel cruzava ervilheiras idênticas em relação a uma determinada característica, durante verias gerações e eliminava sucessivamente aquelas que surgiam com uma variação dessa característica, diferente daquela que ele pretendia obter.
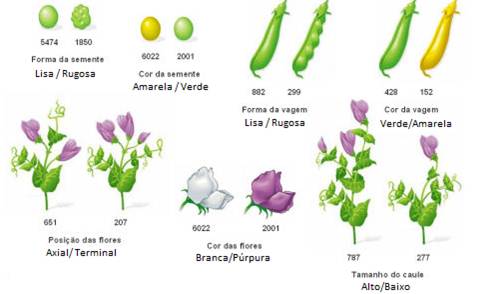
Fig. Características analisadas por Mendel
Após a obtenção de linhas puras, Mendel efectuou cruzamentos parentais, isto é, cruzamentos entre indivíduos de linhas puras mas em que cada indivíduo, envolvido no cruzamento, apresentava uma forma antagónica para a característica em estudo.
Mendel começou por cruzar indivíduos de uma geração parental (vulgarmente designada geração P), que eram linhas puras, mas detentoras de caracteres antagónicos (ex. sementes lisas vs sementes rugosas) – cruzamentos parentais.

Todas as sementes resultantes deste primeiro cruzamento apresentavam o aspecto liso, tendo desaparecido por completo o carácter rugoso. Esta primeira geração filial é designada por F1, geração F1 ou híbridos da primeira geração.
Ao permitir que as sementes da geração F1 germinassem e dessem origem a plantas adultas, que se autopolinizaram, Mendel obteve uma geração F2 onde foram produzidas sementes lisas e rugosas. Mendel calculou a proporção entre as suas classes de sementes e concluiu que a relação era de, aproximadamente, 3:1. Mendel estendeu a sua análise a outros caracteres, tendo obtido um padrão semelhante.
Para todas as características estudadas, Mendel verificou que:
Conclusões de Mendel:
Os estudos desenvolvidos posteriores a Mendel, permitiram verificar que os factores abstractos que Mendel 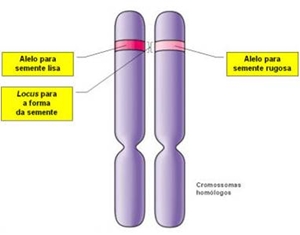 concebeu para explicar a transmissão de características ao longo das gerações existem e correspondem a segmentos de DNA designados genes. Os genes apresentam formas alternativas, sendo cada uma dessas formas chamadas genes alelos ou simplesmente alelos (exemplo: alelo dominante – semente lisa; alelo recessivo – semente rugosas).
concebeu para explicar a transmissão de características ao longo das gerações existem e correspondem a segmentos de DNA designados genes. Os genes apresentam formas alternativas, sendo cada uma dessas formas chamadas genes alelos ou simplesmente alelos (exemplo: alelo dominante – semente lisa; alelo recessivo – semente rugosas).
O local do cromossoma ocupado por um gene, é designado locus, assim, o carácter, por exemplo, “forma da semente” é determinado por dois alelos que se encontram num mesmo local correspondente (locus) em cromossomas homólogos.
Os genes responsáveis por um determinado carácter constituem o genótipo (exemplo: LL, Lr, rr). A expressão de um determinado genótipo é responsável pelo carácter observável ou fenótipo, isto é o fenótipo é as características anatómicas, fisiológicas e bioquímicas que são observáveis, por exemplo, “sementes lisas” e “sementes rugosas”.
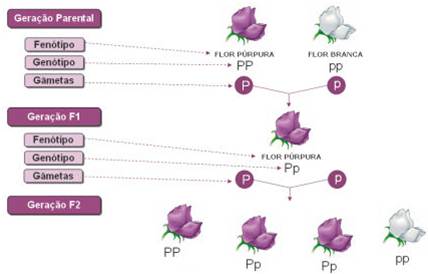
Os resultados de Mendel são actualmente interpretados da seguinte forma:
Para facilitar a visualização das combinações possíveis de gâmetas, recorre-se ao xadrez mendeliano:
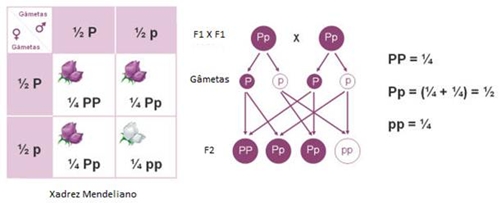
Os indivíduos da geração F1 são 100% heterozigóticos (Pp).
Os indivíduos da geração F2 são:
A análise do xadrez mendeliano permite fazer uma previsão teórica das proporções genotípicas dos indivíduos da geração F2. Na geração F2 podem surgir indivíduos com o mesmo fenótipo (corola púrpura), mas que podem ter dois genótipos diferentes (PP ou Pp), pois o alelo P sendo dominante, e ao estar presente, é o único que se manifesta.
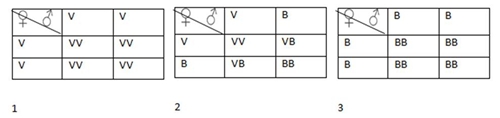
Mendel procurou demonstrar que o fenótipo flores de cor púrpura, poderia aparecer de diferentes genótipos, por isso cruzou algumas ervilheiras com corola púrpura (de genótipo desconhecido) com ervilheiras de corola branca (homozigóticas recessivas).
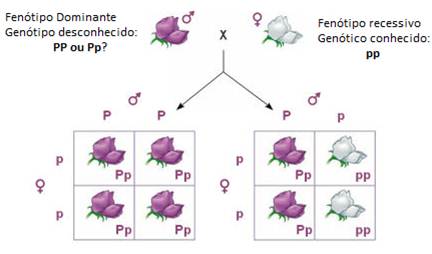
Ao efectuar este tipo de cruzamento é possível descobrir o genótipo do fenótipo dominante.
Nas situações consideradas (imagem), os indivíduos linhas puras recessivos apenas produzem gâmetas portadores do alelo recessivo p. Contudo, os progenitores que apresentam corola púrpura tanto podem ser homozigóticos (PP) como heterozigóticos (Pp).
Assim, no primeiro caso (serem homozigóticos), apenas produzem gâmetas portadores do alelo dominante P e, consequentemente, toda a descendência resultante desse cruzamento (PP X pp) será heterozigótica (Pp) e fenotipicamente uniforme (todos apresentam corola púrpura).
No segundo caso (serem heterozigóticos), os progenitores podem produzir gâmetas portadores do alelo dominante P e gâmetas portadores do alelo recessivo p. Assim, espera-se que 50% dos descendentes deste cruzamento (Pp X pp) sejam heterozigóticos (Pp) e 50% homozigóticos (pp).
Conclui-se, portanto, que a utilização de indivíduos homozigóticos recessivos permite que se manifestem os alelos presentes nos indivíduos que apresentam o fenótipo dominante, e assim, se determine o seu genótipo.
Este tipo de teste só funciona se os resultados da descendência forem numericamente significativos.
Mendel também analisou a transmissão simultânea de duas características – experiências de diibridismo. Ao efectuar o cruzamento de plantas homozigóticas para os dois caracteres em estudo (ex: forma e cor da semente) verificou que:
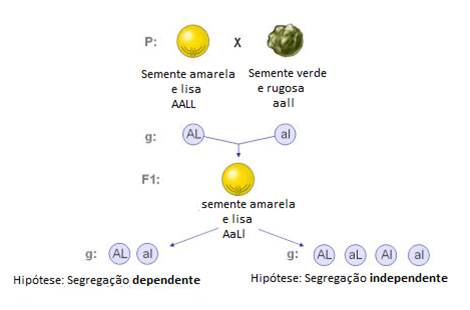
Segregação dependente: os alelos responsáveis pelos dois caracteres em estudo são transmitidos em bloco para os gâmetas, isto é, considerando que os dois caracteres são recebidos dos pais e transmitidos para a descendência sempre juntos (por exemplo: A com L e a com l).
Segregação independente: os alelos responsáveis pelos dois caracteres em estudo são transmitidos de forma independente para os gâmetas, isto é, independentemente para os gâmetas, isto é, independentemente do conjunto de genes que foi herdado de cada progenitor, os indivíduos F1, ao produzirem os seus gâmetas podem combiná-los ao acaso, desde que cada gâmeta receba um alelo para a cor e a forma da semente.
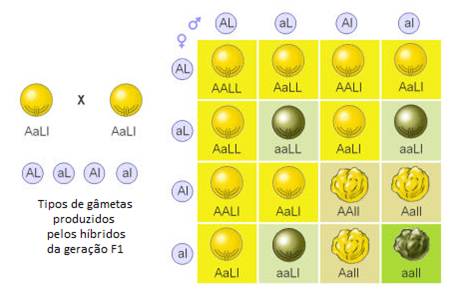
Mendel repetiu estas experiências com diversas combinações diíbridas dos setes caracteres estudados, concluindo que os pares de factores hereditários se comportam de uma forma independente (segregação independente), distribuindo-se ao acaso nos gâmetas.
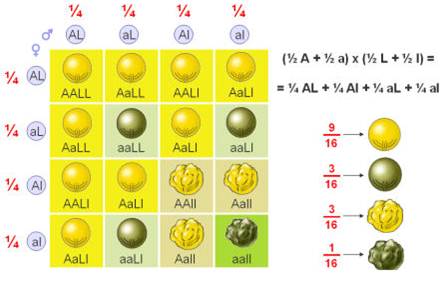
A união aleatória dos 4 tipos de gâmetas origina 16 genótipos, com as seguintes proporções genotípicas:
 9 Amarelas e lisas
9 Amarelas e lisasO facto dos alelos de genes diferentes serem segregados de forma independente durante a formação dos gâmetas é frequentemente designada Segunda Lei de Mendel ou Lei da Segregação Independente.
Existem duas regras básicas das probabilidades com aplicação na transmissão dos caracteres hereditários:
Leis de Mendel actualizadas:
A redescoberta dos trabalhos de Mendel, no inicio do séc. XX, a par do estudo da mitose, da meiose e da fecundação permitiram explicar, através de processos celulares, as conclusões de Mendel:
Alguns estudos sobre a hereditariedade que se realizaram depois de Mendel não respeitavam os princípios mendelianos. Contudo, verificou-se que, mesmo nestas situações, o modo de transmissão das características continuava a ser baseada no “modelo” proposto por Mendel.
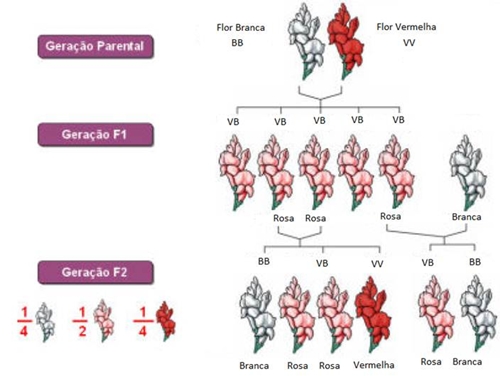
O cruzamento entre as linhas puras (VV x BB) originou uma descendência F1 uniforme, genotipicamente VB e com um fenótipo cor-de-rosa.
A autopolinização de F1, originou 25% de plantas com flores vermelhas, 50% de plantas com flores cor-de-rosa e 25% de plantas com flores brancas, resultantes dos genótipos VV, VB, BB.
Análise da tabela:
Situações em que não há dominância total de um sobre o outro, isto é há a inexistência de uma verdadeira relação dominância/recessividade entre os alelos responsáveis pela cor das flores, ou seja, não há dominância total do alelo vermelho sobre o branco, expressam-se ambos parcialmente (fenótipo intermédio VB). Diz-se, por isso, que existe uma dominância incompleta. Nestes casos, é mais fácil identificar o genótipo, dado que cada fenótipo tem um genótipo diferente.
Outra situação em que não se verifica a relação dominância/recessividade entre os alelos de um determinado gene ocorre quando ambos os alelos se expressam com igual influência na determinação do fenótipo.
Esta situação ocorre, por exemplo, com os genes envolvidos na determinação da cor do pêlo de alguns bovinos.
O cruzamento entre indivíduos de linha pura de cor vermelha (VV) com indivíduos de linha pura de cor branca (BB) origina descendentes que possuem uma mistura de pêlos vermelhos e pêlos brancos, cujo efeito é uma coloração cinzento-avermelhada. Cada um do alelo expressa-se de forma independente pêlo a pêlo. Diz-se que existe uma situação de codominância.
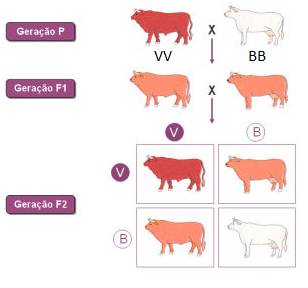
Enquanto, numa situação de dominância incompleta, os indivíduos heterozigóticos apresentam um fenótipo intermédio, resultante do facto de não existir uma dominância completa de um alelo sobre o outro, numa situação de codominância os indivíduos heterozigóticos expressam simultaneamente e de forma independente o fenótipo dos dois homozigóticos.
Nas situações anteriormente apresentadas, cada locus apresentava apenas dois alelos que o poderiam ocupar. Contudo, numa população podem existir mais do que duas formas alélicas concorrentes para um determinado locus. Se existirem três ou mais alelos que podem ocupar os dois loci correspondentes de um par de homólogos, diz-se que esse locus tem alelos múltiplos. Note-se que, mesmo nesta situação, cada individuo possui apenas dois dos diversos alelos possíveis.
PÁGINA 98 (exemplo dos coelhos)
Sendo a relação de dominância → C > Cch > Ch > Ca
Pelo facto de existir uma relação de dominância em sequência entre estes quatro alelos, a sua combinação, dois a dois, permite gerar quatro fenótipos diferentes. Para cada um desse fenótipo existe mais do que um genótipo possível (excepto o coelho albino).
Como exemplo de alelos múltiplos em seres humanos temos os grupos sanguíneos.
Sistema ABO
| Grupo | Genótipo | Aglutinogénios e Anticorpos | Imagem |
| A | AO, AA | Aglutinogénios A+ Anticorpos anti- B |  |
| B | BO, BB | Aglutinogénios B+ Anticorpos anti- A |  |
| O | OO | Sem aglutinogénios + Anticorpos anti- A e anti -B |  |
| AB | AB | Sem aglutinogénios + Sem Anticorpos |  |
Transfusões de sangue
| Doador | Receptor A | Receptor B | Receptor AB | Receptor O |
| A | Sim | Não | Sim | Não |
| B | Não | Sim | Sim | Não |
| AB | Não | Não | Sim | Não |
| O | Sim | Sim | Sim | Sim |
Constata-se que o grupo sanguíneo O, pode doar sangue para os demais grupos sem reacção de incompatibilidade, é denominado doador universal e o AB, por receber sangue dos demais grupos, é denominado receptor universal.
Nas variadas espécies, existem determinados alelos que, quando se reúnem em homozigotia, podem conduzir à morte do seu portador, sendo, por isso, designados alelos letais. Em heterozigotia tais alelos não são letais o que permite a sua manutenção na população. Os seus efeitos podem manifestar-se logo no desenvolvimento embrionário ou após o nascimento.
Cor do pêlo dos gatos:
F1 AA x aa
F2 Aa X Aa

2/3 Cor de pêlo amarelo
1/3 cor de pêlo preto
Os cruzamentos que envolvem genes letais podem conduzir a alterações nas proporções fenotípica observadas em relação às esperadas de acordo com os princípios de Mendel. Este facto resulta da morte de indivíduos portadores do gene letal em homozigotia (no caso AA) antes de completarem o seu desenvolvimento embrionário. Nesta situação, ¼ dos embriões não chega a nascer e , assim, dos sobreviventes. 2/3 terão fenótipo de cor de pêlo amarelo e 1/3 terá a cor do pêlo preto.
Uma forma de evitar a ocorrência de embriões mortos devido à presença dos alelos letais em homozigotia é realizar cruzamentos entre progenitores em que apenas um deles seja portador do gene letal (Aa x aa). Desta forma, irão nascer indivíduos heterozigóticos (Aa) e indivíduos homozigóticos normais (aa).
Nos casos que têm sido apresentados têm sidos explorados até ao momento, o fenótipo resultava da expressão de um único gene. Contudo, são diversas as situações em que dois ou mais genes não alélicos interactuam para produzir uma determinada característica fenotípica de um gene de um segundo locus, isto é, um gene de um determinado locus altera a expressão fenótipica de um gene de um segundo locus. Esta situação é denominada epistasia.
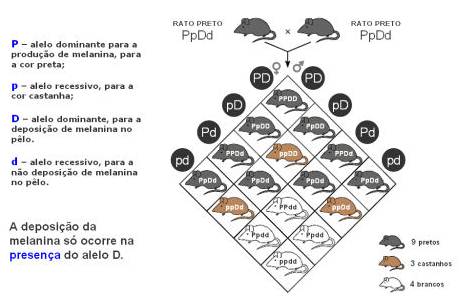
Se considerarmos que se trata de um cruzamento de diibridismo as proporções iriam ser 9:3:3:1, o que não se confirma na experiência.
A presença do alelo P, em homozigotia (PP) ou em heterozigotia (Pp), determinaria a cor preta, enquanto que a cor castanha deveria surgir sempre que o alelo p estivesse em homozigotia (pp). Contudo, a deposição do pigmento que confere a cor do pêlo (preto ou castanho) só ocorre se no locus responsável pela deposição de melanina estiver presente o alelo D, em homozigotia (DD) ou em heterozigotia (Dd). Se neste locus estiverem os dois alelos recessivos (dd), que impedem a deposição do pigmento, o rato será branco, independentemente dos alelos presentes no locus responsável pela cor. Assim, considera-se que o locus responsável pela deposição de melanina é epistático em relação ao locus responsável pela produção deste pigmento, pois o primeiro gene altera a expressão fenotípica do segundo.
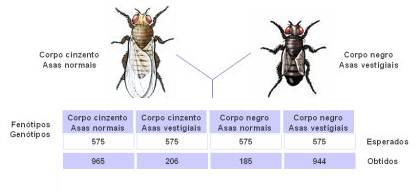
Experiência de Morgan
| Locus responsável pela cor do corpo | Locus responsável pelo tamanho das asas |
| B – alelo responsável pelo corpo cinzento b – alelo responsável pelo corpo negro | V – alelo responsável por asas normais (longas) v- alelo responsável por asas vestigiais |
Geração Parental
Fêmea – Corpo cinzento e asas normais (BBVV). Origina gâmetas BV
Macho – Corpo negro e asas vestigiais (bbvv). Origina gâmetas bv
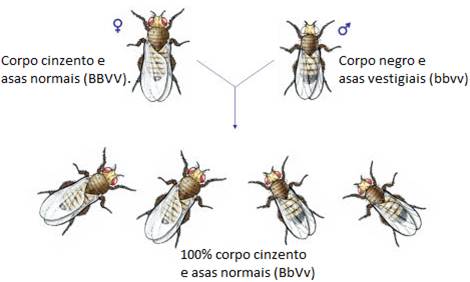
Ao aparecerem 100% Corpo cinzento / asas normais. Verifica-se a 1º Lei de Mendel actualizada porque houve segregação dos genes para os gâmetas. Verifica-se uniformidade na 1ª geração
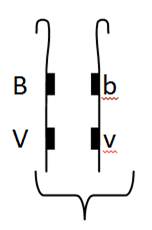 Verifica-se crossing – over entre cromatídeos. Aparecem assim fenótipos recombinantes, ou seja, não aparece na totalidade só dominante ou só recessivo. Não há segregação independente porque os genes estão em bloco. Quanto mais afastados tiverem os genes há maior probabilidade de ocorrer crossing – over.
Verifica-se crossing – over entre cromatídeos. Aparecem assim fenótipos recombinantes, ou seja, não aparece na totalidade só dominante ou só recessivo. Não há segregação independente porque os genes estão em bloco. Quanto mais afastados tiverem os genes há maior probabilidade de ocorrer crossing – over.
Teoricamente só deveria aparecer 2 géneros de fenótipo (BbVv e bbvv) mas apareceram 4 tipos, isto é, embora os loci em questão se encontrem no mesmo cromossoma e por isso tendam a ser transmitidos para os gâmetas em bloco, durante a meiose podem ocorrer fenómenos de recombinação genica devido ao crossing over. Assim embora a maioria dos gâmetas sejam do tipo parental (BV e bv) uma percentagem menor pode apresentar novas combinações dos loci (Bv e bV).
Formaram-se então moscas recombinantes que resultam da fusão de gâmetas recombinantes da F1.
O número de cromossomas do cariótipo de um individuo é incomparavelmente menor do que o número de genes que condicionam o seu fenótipo.
Um cromossoma terá de ter, necessariamente, um grande número de genes.
Os genes dispostos ao longo do mesmo cromossoma dizem-se genes ligados factorialmente ou em linkage e constituem um grupo de ligação factorial, sendo transmitidos em bloco aos descendentes.
Nesta situação não se verifica a segregação independente (2º lei de Mendel) embora possa ser explicada pela teoria cromossómica da hereditariedade.
Como resultado de crossing –over, durante a meiose, os genes podem separar-se e surgir nos gâmetas como se estivessem situados em cromossomas separados.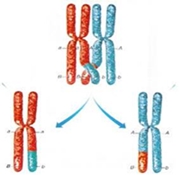
A descendência será qualitativamente igual à prevista numa segregação independente dos genes, mas quantitativamente, surgem em proporções alteradas dado que o crossing-over é meno frequente do que a transmissão em bloco dos genes em caus.
Passado um ano após começar a estudar as Drosophilas, Morgan encontrou um macho de olhos brancos em vez de vermelhos. Estas variações genotípicas são frequentemente designadas fenótipos mutantes (olhos brancos), pelo facto de resultarem de uma mutação. Em oposição, o fenótipo mais comum de um determinado carácter toma a designação de forma ou tipo selvagem (olhos vermelhos).
É costume representar-se a constituição genética das formas alternativas pela letra da inicial da palavra inglesa que expressa a característica que elas manifestam
Alelos para olhos brancos = w (white)
Quando este locus é ocupado pelo alelo da forma selvagem, este representa-se pela mesma letra associada ao sinal +
Alelo para olhos vermelhos = w+
Morgan encontrou um macho mutante de olhos brancos e cruzou-o com uma fêmea de olhos vermelhos. Obteve uma geração F1 com 100% de indivíduos (machos e fêmeas) de olhos vermelhos. Concluiu que o alelo para os olhos vermelhos era dominante sobre o alelo mutante para olhos brancos.
No cruzamento recíproco, entre uma fêmea de olhos brancos e um macho de olhos vermelhos, Morgan obteve resultados diferentes. Obteve uma geração F1 com 50% de indivíduos de olhos vermelhos (todos fêmeas) e 50% de olhos brancos (todos machos).
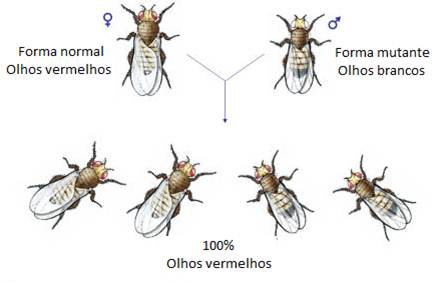
Morgan concluiu que o gene responsável pela cor dos olhos estaria localizado num cromossoma para o qual não existisse um verdadeiro homólogo. Tal verifica-se no par de cromossomas sexuais.
A análise dos resultados das experiências levou Morgan a concluir que o gene responsável pela cor dos olhos estaria localizado no cromossoma X. O gene da cor vermelha não está no cromossoma Y, pois se assim fosse apenas os machos tinham a cor dos olhos vermelhos.
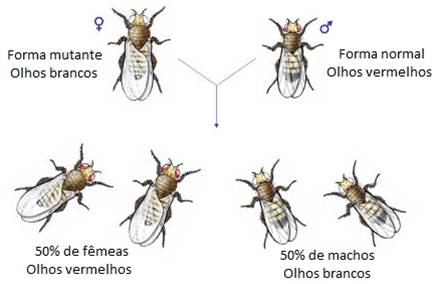
O alelo mutante (responsável pela cor branca dos olhos) é recessivo e, por isso, uma fêmea apenas terá olhos brancos se possuir este alelo em ambos os cromossomas X (o que não se verificou na experiência descrita). A presença do alelo responsável pela cor vermelha, em homozigotia ou em heterozigotia, determinará o aparecimento do fenótipo selvagem, uma vez que é dominante sobre a forma mutante.
Por outro lado, os machos possuem apenas um cromossoma X e, por isso, são portadores apenas de um alelo para as características presentes neste cromossoma. Por esta razão , diz-se que os machos são hemizigóticos para as características determinadas pelos genes localizados no cromossoma X. Assim, a cor dos olhos é determinada apenas por um alelo e, por isso, se os machos herdarem um cromossoma X portador do alelo mutante terão olhos brancos, mas se herdarem um cromossoma X portador do gene normal terão olhos vermelhos.
Nas situações de hemizigotia há uma correspondência directa entre o fenótipo e o genótipo, pois existe apenas um alelo para se manifestar.
Na espécie humana o modo de transmissão da informação genética de geração para geração realiza-se através dos mesmos mecanismos de hereditariedade que operam nas outras espécies.
São diagramas que evidenciam a história da transmissão de um dado carácter ao longo das gerações. A sua analise permite determinar se os genes envolvidos são dominantes ou recessivos ou se estão localizados nos autossomas (transmissão autossómica) ou nos cromossomas sexuais (transmissão ligada aos cromossomas sexuais).
Albinismo
O albinismo é uma doença rara com carácter hereditário que resulta da incapacidade de produzir um pigmento chamado melanina.
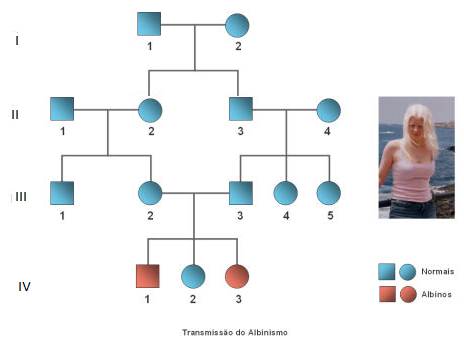
A análise da árvore genealógica permite constatar que progenitores fenotipicamente normais podem originar descendentes de ambos os sexos que manifestam o albinismo. Estes dados permitem inferir que se trata de uma característica autossómica recessiva, isto é, este gene está localizado num autossoma e o alelo mutante é recessivo em relação ao alelo normal.
O alelo do albinismo é um alelo raro. Por esta razão, é mais provável que ele surja em heterozigotia em indivíduos da mesma família, pois têm ancestrais comuns. Assim os cruzamentos consanguíneos, isto é, entre indivíduos aparentados (como o caso dos indivíduos 2 e 3 da 3º Geração), aumentam a probabilidade de o alelo mutante surgir nos descendentes em homozigotia, manifestando-se a anomalia.
Outros dois exemplos deste modo de transmissão são fenilcetonúria (anexos e livro página 112/113) e a surdez.
Algumas anomalias com carácter hereditário podem resultar da expressão de genes dominantes. São exemplos destas anomalias a polidactilia, que se caracteriza pelo aparecimento de mais do que cinco dedos nas mãos e/ou pés. Também a doença de Huntington.
Da análise da transmissão hereditária deste tipo de alelos pode concluir-se que:
Este tipo de transmissões têm o mesmo “funcionamento” que com as moscas Drosophilas.
Hemofilia
A hemofilia é uma doença que resulta da presença de um gene mutante responsável pela síntese de uma proteína necessária para a coagulação sanguínea ocorra. A ausência desta proteína leva a que uma pequena hemorragia possa levar horas até parar, podendo, nalguns casos levar à morte.
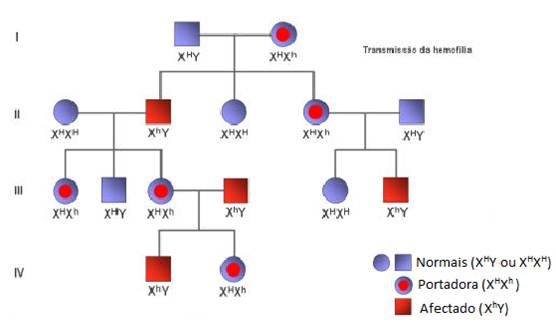
Actualmente, conhece-se mais do que um tipo de hemofilia. Contudo, as duas formas mais frequentes de resultam de genes recessivos localizados no cromossoma X. Considere-se a seguinte simbologia: h- gene responsável pela hemofilia; H- gene normal. Dado que estão localizados no cromossoma X, a representação deverá ser Xh e XH, respectivamente.
À semelhança do que acontece com outras anomalias resultantes da expressão de genes localizados no cromossoma X, verifica-se que afecta quase exclusivamente os homens. Porquê? Uma mulher só será hemofílica se possuir o gene mutante em homozigotia, o que significa que ambos os pais tem de ter um gene com essa doença; além disso admite-se que os gens de hemofílica em homozigotia levam à morte dos embriões.
No caso da Rainha Vitória que passou a doença a toda a sua família, a doença não deixou de existir ao longo das gerações pelo facto de ocorrer um elevado número de cruzamentos consanguíneos entre as famílias reais, o que contribui para a manutenção e manifestação da doença.
Daltonismo
O daltonismo constitui outro exemplo de uma anomalia hereditária resultante de um alelo localizado no cromossoma X. Os indivíduos que manifestam esta anomalia não são capazes de distinguir cores, por exemplo, o verde do vermelho.
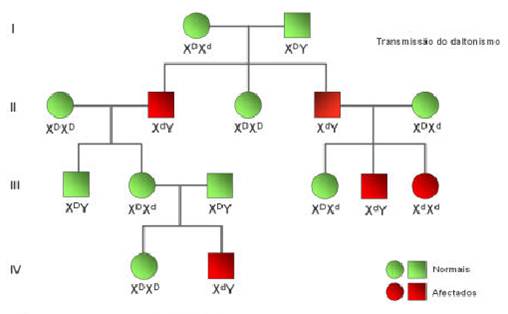
Da análise de árvores genealógicas de fenótipos recessivos ligados ao cromossoma X podemos concluir:
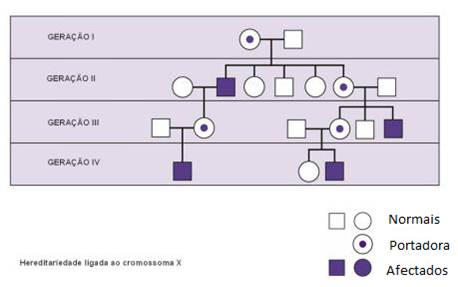
Quando o alelo responsável pela doença se localiza no cromossoma Y todos os filhos de pais com a anomalia apresentam a doença
Revisões (anexos)
Na maioria dos animais, a determinação do sexo envolve apenas diferenças ao nível dos heterossomas (cromossomas sexuais). Contudo, em várias espécies de insectos, a determinação do sexo é feita com base no número total de cromossomas que cada individuo possui. Assim, no caso das abelhas, as fêmeas são diplóides, enquanto que os indivíduos constituídos por células haplóides são do sexo masculino. Nos mamíferos, os indivíduos do sexo feminino possuem dois cromossomas X, e os machos um X e um Y. Nas aves, os machos possuem dois heterossomas iguais (ZZ) e as fêmeas possuem um par ZW.
Determinação genética do sexo em seres humanos (revisões) – página 124 -125
Expressão génica (anexos)
Material genético extra nuclear (livro página 133-134)
Transmissão da informação das mitocôndrias
Alelos: genes que têm a mesma informação contrastante (ex: informação sobre a cor dos olhos – azuis/castanhos) e localizam-se no mesmo locus de cromossomas diferentes.
Linhas Puras: indivíduos que uma vez cruzados entre si, dão sempre origem a descendentes iguais entre si e iguais aos progenitores, em relação à característica considerada.
Híbridos: indivíduos que resultam de um cruzamento parental (ex: LL x rr = Lr)
Monoibridismo: cruzamento entre indivíduos em que se considera a transmissão de um só carácter.
Diibridismo: cruzamento entre indivíduos em que se considera a transmissão de dois caracteres.
Cruzamento Parental: Cruzamento entre indivíduos, de duas linhas puras, que expressam formas antagónicas (diferentes) do carácter considerado.
Homozigóticos: os alelos do par são idênticos e os seus gâmetas são geneticamente semelhantes (ex: PP, pp). Nota: os indivíduos de linhas puras, são sempre homozigóticos, mas nem todos os homozigóticos são de linhas puras.
Heterozigóticos: os alelos do par são diferentes e os seus gâmetas portadores de um alelo ou de outro (ex: Pp). Nota: Todos os híbridos são heterozigóticos, mas nem todos os heterozigóticos são híbridos.
Fenótipo: características observáveis (morfológicas ou funcionais) do indivíduo, resultante da expressão do genótipo (em interacção com o ambiente)
Genótipo: constituição genética de um indivíduo em relação à(s) características consideradas
O genoma de um individuo pode sofrer alterações designadas mutações.
As mutações podem ocorrer a nível das células somáticas ou a nível dos gâmetas. No primeiro caso, a mutação terá reflexo no conjunto de células descendentes daquela que sofreu a mutação, podendo, por isso, restringir-se a uma pequena zona do organismo. Estas mutações somáticas são responsáveis por alguns tipos de cancros. Se a mutação ocorrer ao nível dos gâmetas, todas as células que eventualmente descendam desse gâmeta serão portadores da mutação, isto é, a mutação passará à geração seguinte.
(imagem da página 145)
Quando as mutações envolvem apenas um gene (ou um número muito restrito de genes), tomam a designação de mutações genicas. Contudo, as alterações podem ser mais extensas, envolvendo a estrutura ou mesmo o número de cromossomas, nesse caso designam-se mutações cromossómicas.
(Ler página 146-148)
Uma alteração ao nível dos genes deve-se à substituição de nucleótidos (como a anemia falciforma), que podem ser:

Noutros casos, a molécula de DNA perde (delecção) ou ganha (inserção) um nucleótido, alterando-se por completo a mensagem a partir do codão onde ocorreu a mutação.
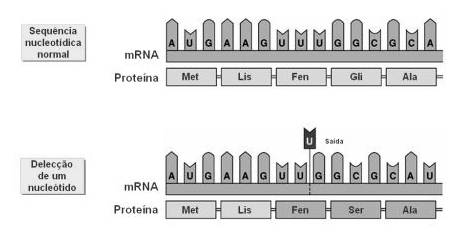
Mutação por delecção
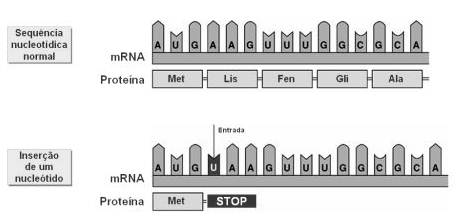
Mutação por inserção
Durante a meiose, verificam-se fenómenos de crossing-over, que permitem a troca de material genético entre cromossomas homólogos, contribuindo para o aumento da variabilidade genética. Contudo, por vezes, durante este complexo processo, ocorrem erros que se traduzem pela alteração da estrutura ou do número de cromossomas.
Nas mutações cromossómicas estruturais, verifica-se a manutenção do número de cromossomas, alterando-se apenas o arranjo e/ou números de genes.
Este tipo de mutações pode ser de:
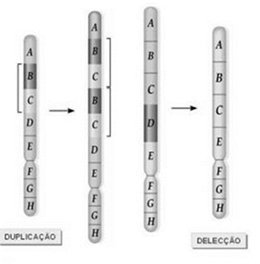
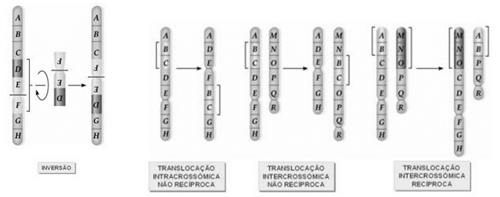
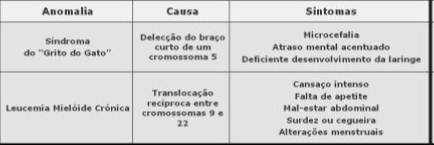
Exemplos de mutações cromossómicas estruturais:
Um cariótipo que apresente o número normal de cromossomas diz-se euplóide. Normalmente, os organismos são haplóides (n) ou diplóides (2n). Contudo, erros que podem ocorrer durante a meiose podem multiplicar o conjunto de todos os cromossomas, originando poliploidias. Em outras situações, os erros de separação dos cromossomas envolvem apenas um determinado par de homólogos, originando aneuploidias.

Durante a meiose pode ocorrer quer uma não – disjunção de cromossomas homólogos na divisão I, quer uma não disjunção de cromatídeos na divisão II. De ambos os casos resultam células com excesso ou com défice de cromossomas. A não disjunção pode envolver cromossomas autossómicos como os cromossomas sexuais. Na espécie humana são raras as aneuploidias que envolvem autossomas e que sejam compatíveis com a vida, sendo que acontecem abortos.
Lê – página 163-165

(depois no livro explica a formação de cancro! Como é tudo importante é mais fácil leres J - página 153-158)
(para poupar papel e tinta, ler página 166-175)
 Angola
Angola Moçambique
Moçambique Cabo Verde
Cabo Verde Brasil
Brasil Inglês
Inglês Portugal
Portugal