
O teu país
Lorem ipsum dolor sit amet, consectetur adipiscing elit, sed do eiusmod
Lorem ipsum dolor sit amet, consectetur adipiscing elit, sed do eiusmod

Todos os trabalhos publicados foram gentilmente enviados por estudantes – se também quiseres contribuir para apoiar o nosso portal faz como o(a) Andreia Lopes e envia também os teus trabalhos, resumos e apontamentos para o nosso mail: geral@notapositiva.com.
Resumo/Apontamentos escolares sobre o Património Genético, realizado no âmbito da disciplina de Biologia (12º ano)...
A genética é uma ciência jovem, iniciada nos primórdios do século XX, altura em que foram redescobertas as leis da hereditariedade, estabelecidas por Gregor Mendel em 1865.
A genética, área do conhecimento que tem como objecto de estudo os genes, procura respostas para a forma como se processa a herança genética, tendo-se tornado um assunto central na Biologia.
As leis de Mendel foram estabelecidas a partir da análise estatística de resultados experimentais obtidos em cruzamentos realizados com a ervilheira (Pisum sativum). A ervilheira é uma planta que apresenta características adequadas ao estudo da hereditariedade, uma vez que possui características diferenciadas e contrastantes, fácil cultivo e rápido crescimento, obtenção de várias gerações e elevado número de descendentes num curto período de tempo e flor com estrutura adequada à auto – polinização e ao controlo da fecundação.
Mendel começou a sua investigação com a análise da transmissão de um carácter isoladamente (monibridismo).
O monibridismo é o cruzamento entre indivíduos em que se considera a transmissão de um só carácter. Para cada uma das características seleccionadas e durante dois anos, Mendel tentou isolar linhas puras, ou seja, indivíduos que, cruzados entre si, originam sempre descendência toda igual entre si e igual aos progenitores, para o carácter considerado.

Seleccionadas as linhas puras, Mendel efectuou cruzamentos parentais, isto é, cruzamentos entre indivíduos pertencentes a essas linhas puras, que expressam de forma antagónica o carácter considerado. Para conseguir este cruzamento, Gregor Mendel recorreu ao modo de polinização cruzada artificial. A análise dos dados recolhidos nos ensaios revelou que:
Para simplificar o tratamento dos dados utiliza-se símbolos para representar os factores. Assim L designa o alelo responsável pela forma dominante e l o alelo responsável pela forma recessiva das sementes da ervilheira.
Assim, no exemplo das ervilheiras, partimos de uma geração parental onde os progenitores são homozigóticos (LL e ll) e fenotipicamente, um origina sementes lisas e o outro rugosas. A geração F1 é formada por indivíduos todos heterozigóticos (Ll) e fenotipicamente com sementes lisas. Estes indivíduos produzem dois tipos de gâmetas: gâmetas que transportam o alelo dominante (L) e gâmetas que transportam o alelo recessivo (l). estes gâmetas vão combinar-se de todas as formas possíveis para originar a 2ª geração (F2).
Os cruzamentos costumam ser representados através de um quadro ou matriz de cruzamento com duas entradas. Numa entrada do quadro inscreve-se a constituição genética dos tipos de gâmetas possíveis para um dos progenitores; numa outra entrada, o mesmo tipo de informação para o outro progenitor.

Uma das maneiras de averiguar o genótipo de indivíduos que fenotipicamente revelam a característica do alelo dominante é cruzar esses mesmos indivíduos com indivíduos homozigóticos recessivos e analisar a respectiva descendência. A estes cruzamentos dá-se o nome de cruzamentos – teste ou retrocruzamentos. Estes funcionam para descendências numericamente significativas.
Mendel também analisou a transmissão simultânea de duas características – experiências de diibridismo. Ao efectuar cruzamentos de plantas homozigóticas para os dois caracteres em estudo (ex.: forma e cor da semente) verificou que:

Mendel conclui que os pares de factores hereditários se comportam de uma forma independente, distribuindo-se ao acaso nos gâmetas. A união aleatória dos quatro tipos de gâmetas origina dezasseis fenótipos, com as seguintes proporções fenotípicas:

Uma determinada espécie de planta origina flores de cor branca, vermelhas e rosa. Nos cruzamentos  efectuados, os indivíduos da geração F1, resultantes do cruzamento parental, apresentam todos flores cor-de-rosa, que corresponde a um terceiro fenótipo, intermédio entre o vermelho e o branco. Trata-se de um caso de dominância incompleta, ou seja, situações em que não há dominância total de um alelo sobre o outro, isto é, não existe dominância nem recessividade. Expressam-se ambos parcialmente pelo que, em relação ao gene considerado, surge um fenótipo intermédio em indivíduos heterozigóticos.
efectuados, os indivíduos da geração F1, resultantes do cruzamento parental, apresentam todos flores cor-de-rosa, que corresponde a um terceiro fenótipo, intermédio entre o vermelho e o branco. Trata-se de um caso de dominância incompleta, ou seja, situações em que não há dominância total de um alelo sobre o outro, isto é, não existe dominância nem recessividade. Expressam-se ambos parcialmente pelo que, em relação ao gene considerado, surge um fenótipo intermédio em indivíduos heterozigóticos.
Existem ainda outras situações em que ocorre co-dominância, ou seja, situações em que os alelos de um par de genes se expressam ambos completamente no fenótipo, em indivíduos heterozigóticos.
| Relações entre alelos | Genótipos | Fenótipos |
| Dominância e recessividade |
|
|
| Dominância incompleta |
|
|
| Co-dominância |
|
|
A redescoberta dos trabalhos de Mendel, no início do século XX, a par do estudo da mitose, da meiose e da fecundação permitiram a Sutton e Boveri (1902) explicar, através de processos celulares, as conclusões de Mendel (teoria cromossómica da hereditariedade):
Thomas H. Morgan foi um embriologista da Universidade da Colúmbia nos EUA. Desenvolveu, a partir de 1910, estudos genéticos com a mosca – da – fruta (Drosophila melanogaster). Em 1933 recebeu o Prémio Nobel.
A forma de Drosophila predominante na Natureza tem o corpo cinzento, olhos vermelhos e asas longas e denomina-se forma selvagem. Outras formas alternativas (mutantes) podem revelar o corpo negro, olhos brancos ou asas vestigiais, entre outras características.

É costume representar-se a constituição genética das formas alternativas pela letra inicial da palavra inglesa que expressa a característica que elas manifestam.
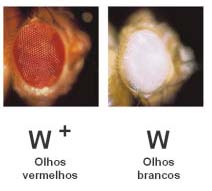 Alelo para olhos brancos – w (de white)
Alelo para olhos brancos – w (de white)
Quando este locus é ocupado pelo alelo da forma selvagem, este representa-se pela mesma letra associada ao sinal +.
Alelo para olhos vermelhos – w+
Morgan encontrou um macho mutante de olhos brancos e cruzou-o com uma fêmea de olhos vermelhos. Obteve uma geração F1 com 100% de indivíduos (machos e fêmeas) de olhos vermelhos. Concluiu que o alelo para olhos vermelhos era dominante sobre o alelo mutante para olhos brancos.

No cruzamento recíproco, entre uma fêmea de olhos brancos e um macho de olhos vermelhos, Morgan obteve resultados diferentes. Obteve uma geração F1 com 50% de indivíduos de olhos vermelhos (todos fêmeas) e 50% de olhos brancos (todos machos). Morgan conclui que o gene responsável pela cor dos olhos estaria localizado num cromossoma para o qual não existisse um verdadeiro homólogo. Tal verifica-se no par de cromossomas sexuais.

As fêmeas possuem dois cromossomas X (verdadeiros homólogos), os machos possuem um cromossoma X e um cromossoma Y, sem genes correspondentes (não são verdadeiros homólogos). As fêmeas designam-se homogaméticas e os machos heterogaméticos.
Morgan concluiu que o gene responsável pela cor dos olhos estaria no cromossoma X. os machos manifestam o único alelo, localizado no cromossoma X (denominando-se hemizigóticos).
As características hereditárias que dependem de genes localizados no cromossoma X dizem-se características ligadas ao sexo.
O número de cromossomas do cariótipo de um indivíduo é incomparavelmente menor do que o número de genes que condicionam o seu fenótipo. Um cromossoma terá de ter, necessariamente, um grande número de genes. Os genes dispostos ao longo do mesmo cromossoma dizem-se genes ligados factorialmente ou em  linkage e constituem um grupo de ligação factorial, sendo transmitidos em blocos aos descendentes.
linkage e constituem um grupo de ligação factorial, sendo transmitidos em blocos aos descendentes.
Nesta situação não se verifica a segregação independente (2ª lei de Mendel) embora possa ser explicada pela teoria cromossómica da hereditariedade.
No caso considerado cruzaram-se indivíduos de duas linhas puras com características antagónicas. Os diíbridos da 1ª geração são uniformes, manifestando as características determinadas pelos alelos dominantes (corpo cinzento e asas longas). O genótipo é heterozigótico em relação aos dois pares de genes, b+bvg+vg. Na geração F2 surgem 50% de indivíduos de corpo cinzento a asas longas e 50% de indivíduos de corpo negro e asas vestigiais.
Com estes resultados conclui-se que o híbrido apenas produziu dois tipos de gâmetas: um tipo portador dos alelos b+vg+ e outro tipo portador dos alelos bvg. Assim conclui-se também que os genes que se situam no mesmo cromossoma (ligados factorialmente) transmitem-se à descendência em conjunto, não ocorrendo segregação independente. Mas nem sempre isto acontece. Como resultado de crossing-over, durante a meiose, os genes podem separar-se e surgir nos gâmetas como se estivessem situados em cromossomas separados.
A descendência será qualitativamente igual à prevista numa segregação independente dos genes; quantitativamente, surgem em proporções alteradas dado que o crossing-over é menos frequente do que a transmissão em bloco dos genes em causa.

 Angola
Angola Moçambique
Moçambique Cabo Verde
Cabo Verde Brasil
Brasil Inglês
Inglês Portugal
Portugal